The topic of enzyme catalysis, because virtually all cellular reactions or processes are mediated by protein (or, in certain cases, RNA) catalysts called enzymes. It is only as we explore the nature of enzymes and their catalytic properties that we begin to understand how reactions that are energetically feasible actually take place in cells and how the rates of such reactions are controlled.
Consider why thermo-dynamically spontaneous reactions do not usually occur at appreciable rates without a catalyst, and then we will look at the role of enzymes as specific biological catalysts, the rate of an enzyme-catalysed reaction is affected by the concentration of substrate available to it, as well as some of the ways in which reaction rates are regulated to meet the needs of the cell.

Activation Energy and the Metastable State are familiar with many reactions that are thermodynamically feasible yet do not occur to any appreciable extent. An obvious example is the oxidation of glucose. This reaction (or series of reactions, really) is highly exergonic (�G°’ = -686 kcal/mol) and yet does not take place on its own. In fact, glucose crystals or a glucose solution can be exposed to the oxygen in the air indefinitely, and little or no oxidation will occur. The cellulose in the paper on which these words are printed is another example and so, for that matter, are consisting as you do of a complex collection of thermodynamically unstable molecules.
Not nearly as familiar, but equally important to cellular chemistry, are the many thermodynamically feasible reactions in cells that could go, but do not proceed at an appreciable rate on their own. As an example, consider the high-energy molecule adenosine triphosphate (ATP), which has a highly favourable �G°’(-7.3 kcal/mol) for the hydrolysis of its terminal phosphate group to form the corresponding diphosphate (ADP) and inorganic phos-phate (Pi): ATP + H2O ADP + Pi This reaction is very exergonic under standard conditions and is even more so under the conditions that prevail in cells. Yet despite the highly favorable free energy change, this reaction occurs only slowly on its own, so that ATP remains stable for several days when dissolved in pure water.
This property turns out to be shared by many biologically important molecules and reactions, and it is important to understand why.
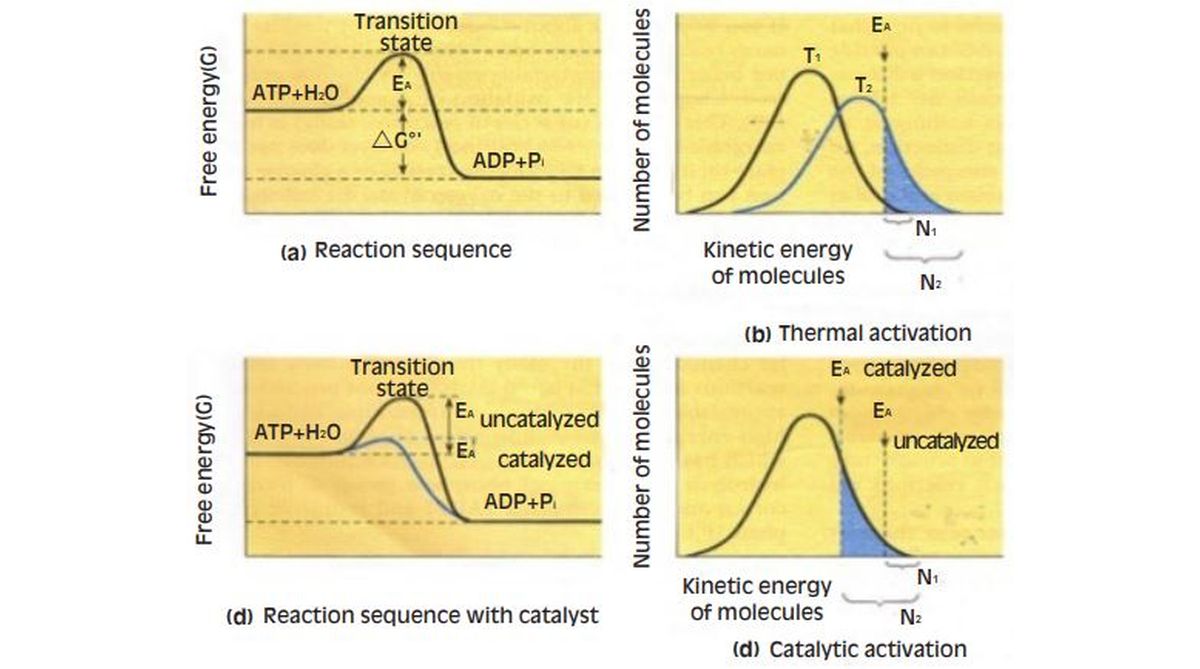
Before a chemical reaction can occur, the activation energy barrier must be overcome. Molecules that should react with one another often do not because they lack sufficient energy. For > every reaction, there is a specific activation energy (EA), which is the minimum amount of energy that reactants must have before collisions between them will be successful in giving rise to products. More specifically, reactants need to reach an intermediate chemical stage called the transition state, the free energy of which is higher than that of the initial reactants.
The actual rate of a reaction is always proportional to the fraction of molecules that have energy content equal to or greater than EA. When in solution at room temperature, molecules of ATP and water move about readily, each possessing a certain amount of energy at any instant. The energy distribution among molecules will be bell-shaped; some molecules will have very little energy, some will have a lot, and most will be near average. The important point is that the only molecules that are capable of reacting at a given instant are those with enough energy to exceed the activation energy barrier.
The metastable state is a result of the activation barrier for most biologically important reactions at normal cellular temperatures, the activation energy is sufficiently high that the proportion of molecules possessing that much energy at any instant is extremely small. Accordingly, the rates of uncatalysed reactions in cells are very low, and most molecules appear to be stable even though they are potential reactants in thermodynamically favoured reactions. Such seemingly stable molecules are said to be in a metastable state. For cells and cell biologists, high activation energies and the resulting metastable state of cellular constituents are crucial, because life by its very nature is a system maintained in a steady state a long way from equilibrium. Otherwise, all reactions would proceed quickly to equilibrium, and life would be impossible.
Catalysts overcome the activation energy barrier as it is to the maintenance of the metastable state, the activation energy requirement is a barrier that must be overcome if desirable reactions are to proceed at reasonable rates. Since the energy content of a given molecule must exceed EA before that molecule is capable of undergoing reaction, the only way a reaction involving metastable reactants can be made to proceed at an appreciable rate is to increase the proportion of molecules with sufficient energy. This can be achieved either by increasing the average energy content of all molecules or by lowering the activation energy requirement. One way to increase the energy content of the system is by the input of heat. Simply increasing the temperature of the system from T, to T2 will increase the kinetic energy of the average molecule, thereby ensuring a greater number of reactive molecules (N2 instead of N,). Thus, the hydrolysis of ATP could be facilitated by heating the solution, giving each ATP and water molecule more energy.
The problem with using an elevated temperature is that such an approach is incompatible with life, because biological systems require a relatively constant temperature. Cells are basically isothermal (constanttemperature) systems and require isothermal methods to solve the activation problem.
Providing a reactive surface is the task of a catalyst, an agent that enhances the rate of a reaction by lowering the energy of activation, thereby ensuring that a higher proportion of the molecules are energetic enough to undergo reaction without the input of heat. A primary feature of a catalyst is that it is not permanently changed or consumed as the reaction proceeds. It simply provides a suitable surface and environment to facilitate the reaction.
For a specific example of catalysis, consider the decomposition of hydrogen peroxide (H2O2) into water and oxygen: 2H2O2 2 H2O + O2 This is a thermodynamically favoured reaction, yet hydrogen peroxide exists in a metastable state because of the high activation energy of the reaction. However, if we add a small number of ferric ions (Fe3+) to a hydrogen per-oxide solution, the decomposition reaction proceeds about 30,000 times faster than without the ferric ions. Clearly, Fe3+ is a catalyst for this reaction, lowering the activation energy and thereby ensuring that a significantly greater proportion (30,000-fold more) of the hydrogen peroxide molecules possess adequate energy to decompose at the existing temperature without the input of added energy.
In cells, the solution to hydrogen peroxide breakdown is not the addition of ferric ions but the enzyme catalyse, an iron-containing protein.
In the presence of catalyse, the reaction proceeds about 100,000,000 times faster than the uncatalysed reaction.
Catalyse contains iron atoms bound in chemical structures called porphyrins, thus taking advantage of inorganic catalysis within the context of a protein molecule. This combination is obviously a much more effective catalyst for hydrogen peroxide decomposition than ferric ions by themselves.
The rate enhancement of about 108 for catalyse is not at all an atypical value; the rate enhancements of enzyme-catalysed reactions range from 107 to as high as 1014 compared with the uncatalysed reaction. These values underscore the extraordinary importance of enzymes as catalysts and bring us to the main theme.
(The writer is associate professor and head, department of botany, Ananda Mohan College, Kolkata)